
B клетки памяти где образуются
Роль В-лимфоцитов в презентации антигена
В-клетки способны поглощать свои мембранные иммуноглобулины вместе со связанным ими антигеном, а затем презентировать фрагменты антигена в комплексе с молекулами МНС класса II. При низкой концентрации антигена и при вторичном иммунном ответе В-клетки могут выполнять роль основных антигенпрезентирующих клеток.
B-лимфоци́ты (B-клетки, от bursa fabricii птиц, где впервые были обнаружены) — функциональный тип лимфоцитов, играющих важную роль в обеспечении гуморального иммунитета.
У эмбрионов человека и других млекопитающих B-лимфоциты образуются в печени и костном мозге из стволовых клеток, а у взрослых млекопитающих — в красном костном мозге.
При контакте с антигеном или стимуляции со стороны T-клеток некоторые B-лимфоциты трансформируются в плазматические клетки, способные к продукции антител. Другие активированные B-лимфоциты превращаются в B-клетки памяти.
Маркеры В-клеток
Характерной особенностью В-клеток является наличие поверхностных мембрано-связанных антител, относящихся к классам IgM и IgD. В комплексе с с другими поверхностными молекулами иммуноглобулины формируют антиген-распознающий рецептивный комплекс, ответственный за узнавание антигена. Также на поверхности В-лимфоцитов расположены антигены МНС класса II, важные в кооперации с Т-клетками, также на некоторых клонах В- лимфоцитов присутствует CD5 маркер, общий с Т-клетками. Рецепторы C3b компонента комплемента(Cr1, CD35) и C3d (Cr2,CD21) имеют определённую роль в активации В-клеток. Следует отметить, что маркеры CD19, CD20 и CD22 используются для идентификации В-лимфоцитов. Также на поверхности В-лимфоцитов обнаружены Fc-рецепторы.
Содержание
В-клетки
Экспрессия всеми В-клетками мембранных иммуноглобулинов позволяет осуществляться клональному отбору под действием антигена. При созревании, антиген-стимулировании и пролиферации существенно меняется набор маркеров В-клеток. По мере созревания В-клетки переключаются от синтеза IgM и IgD на синтез IgG, IgA, IgE (при этом у клеток сохраняется способность синтезировать также IgM и IgD- вплоть до трёх классов одновременно). При переключении синтеза изотипов антигенная специфичность антител сохраняется. Различают:
- Собственно В-клетки (ещё называемые «наивными» В-лимфоцитами)- неактивированные В-лимфоциты, не пконтактировавшие с антигеном. Не содержат тельца Голла, в цитоплазме рассеяны монорибосомы. Полиспецифичны и имеют слабое сродство к многим антигенам.
- В-клетки памяти - активированые В-лимфоциты, посредством кооперации с Т-клетками вновь перешедшие в стадию малых лимфоцитов. Являются долгоживущим клоном В-клеток, обеспечивая быстрый иммунный ответ и выработку большого количества иммуноглобулинов при повторном введении того же антигена. Названы клетками памяти, так как позволяют иммунной системе «помнить» антиген на протяжении многих лет после прекращения действия антигена. В-клетки памяти обеспечивают долговременный иммунитет.
- Плазматические клетки- являются последним этапом дифференцировки активированных В-клеток, провзаимодействовавшими с антигеном. В отличие от остальных В-клеток несут мало мембранных антител и способны секретировать растворимые антитела. Являются большими клетками с эксцентрично расположенным ядром и развитым синтетическим аппаратом- шероховатый эндоплазматический ретикулум занимает почти всю цитоплазму, также развит и аппарат Гольджи. Являются короткоживущими клетками (2-3 дня) и быстро элиминируются при отсутствии антигена, вызвавшего иммунный ответ.
Активация В-клеток
Антиген-презентирующая клетка (макрофаги, клетки Купфера, фолликулярные дендритные клетки, интердигитальные дендритные клетки и т. д.) вскоре после переваривания патогена выносит эпитопы на поверхность клетки при помощи МНС I или II (в зависимости от природы антигена), делая их доступными для Т-клеток. Т-хелпер при помощи Т-клеточного рецептора распознаёт комплекс эпитоп-МНС. Активированный Т-хелпер выделяет цитокины, усиливающие антиген-презентирующую функцию, а также цитокины, активирующих В-лимфоцит - индукторы активации и пролиферации. В-лимфоциты присоединяются при помощи мембрано-связанных антител, выступающих в роли рецепторов, к "своему" антигену и в зависимости от получаемых от Т-хелпера сигналов пролиферируют и дифференцируются в плазматическую клетку, синтезирующую антитела, либо перерождается в В-клетку памяти. При этом от качества и количества антигена будет зависеть исход данной трёхклеточной системы взаимодействия. Данный механизм справедлив для полипептидных антигенов, относительно неустойчивых к фагоцитарному процессингу - т. н. тимус-зависимых антигенов. Для тимус-независимых антигенов (обладающих высокой полимерностью с часто повторяющимися эпитопами, относительно устойчивых к фагоцитарному перевариванию и обладающих свойствами митогена) участия Т-хелпера не требуется - активация В-лимфоцитов происходит по тимус-независимому пути, В-лимфоциты связываются с данными антигенами, а за счёт их собственной митогенной активности будет происходить пролиферация В-лимфоцитов и активация.
В-клетки
Экспрессия всеми В-клетками мембранных иммуноглобулинов позволяет осуществляться клональному отбору под действием антигена. При созревании, антиген-стимулировании и пролиферации существенно меняется набор маркеров В-клеток. По мере созревания В-клетки переключаются от синтеза IgM и IgD на синтез IgG, IgA, IgE (при этом у клеток сохраняется способность синтезировать также IgM и IgD- вплоть до трёх классов одновременно). При переключении синтеза изотипов антигенная специфичность антител сохраняется. Различают:
- Собственно В-клетки (ещё называемые «наивными» В-лимфоцитами)- неактивированные В-лимфоциты, не пконтактировавшие с антигеном. Не содержат тельца Голла, в цитоплазме рассеяны монорибосомы. Полиспецифичны и имеют слабое сродство к многим антигенам.
- В-клетки памяти - активированые В-лимфоциты, посредством кооперации с Т-клетками вновь перешедшие в стадию малых лимфоцитов. Являются долгоживущим клоном В-клеток, обеспечивая быстрый иммунный ответ и выработку большого количества иммуноглобулинов при повторном введении того же антигена. Названы клетками памяти, так как позволяют иммунной системе «помнить» антиген на протяжении многих лет после прекращения действия антигена. В-клетки памяти обеспечивают долговременный иммунитет.
- Плазматические клетки- являются последним этапом дифференцировки активированных В-клеток, провзаимодействовавшими с антигеном. В отличие от остальных В-клеток несут мало мембранных антител и способны секретировать растворимые антитела. Являются большими клетками с эксцентрично расположенным ядром и развитым синтетическим аппаратом- шероховатый эндоплазматический ретикулум занимает почти всю цитоплазму, также развит и аппарат Гольджи. Являются короткоживущими клетками (2-3 дня) и быстро элиминируются при отсутствии антигена, вызвавшего иммунный ответ.
Роль В-лимфоцитов в презентации антигена
В-клетки способны интернализовать свои мембранные иммуноглобулины вместе со связанным с ними антигеном и затем презентировать фрагменты антигена в комплексе с молекулами МНС класса II. При низкой концентрации антигена и при вторичном иммунном ответе В-клетки могут выполнять роль основных антигенпрезентирующих клеток.
Дифференциация В-лимфоцитов
Лимфоциты происходят от плюрипотентных стволовых клеток, дающих также начало всем клеткам крови. Дифференциация стволовых клеток крови по эритроидному, миелоидному либо лимфоидному пути зависит от микроокружения (в случае птиц дифференциация стволовых клеток в В-лимфоциты происходит в фабрициевой сумке, у млекопитающих в костном мозге, где также происходит дифференциация по миелоидному и эритроидному пути). Дифференциация В-лимфоцитов условно делится на две стадии — антигеннезависимую (в которую происходит перестройка генов иммуноглобулинов и их экспрессия) и антигензависимую (при которой происходит активация, пролиферация и дифференциация в плазматические клетки).
- Пре-В-Клетки-предшественники не синтезируют тяжёлых и лёгких цепей, содержат зародышевые H и L гены, но содержат антигенный маркер, общий с зрелыми пре-В-клетками.
- Ранние пре-В-клетки — D-J перестройки в Н генах.
- Поздние пре-В-клетки — V-DJ перестройки в Н генах.
- Большие пре-В-клетки Н-гены VDJ-перестроены; в цитоплазме имеются тяжёлые цепи класса μ.
- Малые пре-В-клетки — V-J перестройки в L генах; в цитоплазме имеются тяжёлые цепи класса μ.
- Малые незрелые В-клетки — L гены VJ-перестроены; синтезируют Н и L- цепи; на мембране расположены иммуноглобулины.
- Зрелые В-клетки- начало синтеза IgD.
В-клетки клетки поступают из костного мозга во вторичные лимфоидные органы (селезёнку и лимфатические узлы), где происходит их дальнейшее созревание, антиген-презентирование, пролиферация и дифференциация в плазматические клетки и В-клетки памяти.
Созревание В-лимфоцитов
Из костного мозга пре-В-клетки мигрируют в тимуснезависимые зоны лимфоидных органов. Так, в физиологических условиях в селезёнке В-лимфоциты располагаются в краевой зоне белой пульпы, в лимфатических узлах — в наружной зоне кортикального слоя, где они формируют зародышевые центры фолликулов. Сигналы, определяющие судьбу и дифференцировку этих иммунокомпетентных клеток, поступают из красного костного мозга, стромальных клеток и других клеток иммунной системы.
На периферии (вне костного мозга) В-лимфоциты приобретают характерные для них поверхностно-клеточные маркёры. Продолжительность жизни В-лимфоцитов различна — от многих лет (В-клетки памяти) до нескольких недель (клоны плазматических клеток).
После антигенной стимуляции В-лимфоциты дифференцируются в плазматические клетки (интенсивно синтезирующие и секретирующие AT) и В-клетки памяти. Плазматические клетки синтезируют Ig того же класса, что и мембранный Ig В-лимфоцита-предшественника.
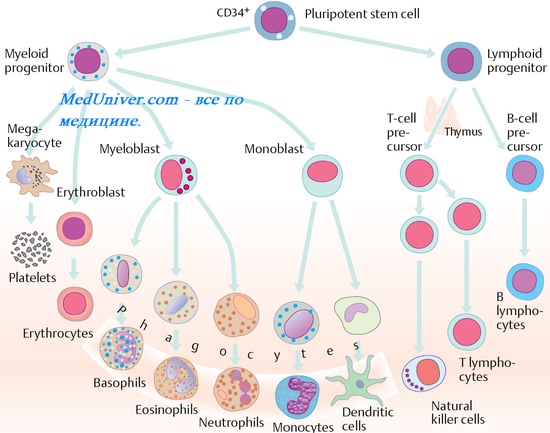
Дифференцировка В-лимфоцитов
Лимфоциты происходят от полипотентных стволовых клеток, дающих также начало всем клеткам крови. Дифференцировка стволовых клеток крови по эритроидному, миелоидному либо лимфоидному пути зависит от микроокружения (в случае птиц дифференцировка стволовых клеток в В-лимфоциты происходит в фабрициевой сумке, у млекопитающих в костном мозге, где также происходит дифференцировка по миелоидному и эритроидному пути). Дифференцировка В-лимфоцитов условно делится на две стадии — антигеннезависимую (в которую происходит перестройка генов иммуноглобулинов и их экспрессия) и антигензависимую (при которой происходит активация, пролиферация и дифференцировка в плазматические клетки). Выделяют следующие промежуточные формы созревающих В-лимфоцитов:
- Ранние предшественники В-клеток — не синтезируют тяжёлых и лёгких цепей иммуноглобулинов, содержат зародышевые IgH и IgL гены, но содержат антигенный маркер, общий со зрелыми пре-В-клетками.
- Ранние про-В-клетки — D-J перестройки в IgН генах.
- Поздние про-В-клетки — V-DJ перестройки в IgН генах.
- Большие пре-В-клетки — IgН гены VDJ-перестроены; в цитоплазме имеются тяжёлые цепи класса μ, экспрессируется пре-В-клеточный рецептор.
- Малые пре-В-клетки — V-J перестройки в IgL генах; в цитоплазме имеются тяжёлые цепи класса μ, экспрессируется пре-В-клеточный рецептор.
- Малые незрелые В-клетки — IgL гены VJ-перестроены; синтезируют тяжёлые и лёгкие цепи; на мембране экспрессируются иммуноглобулины.
- Зрелые В-клетки — начало синтеза IgD.
В-клетки поступают из костного мозга во вторичные лимфоидные органы (селезёнку и лимфатические узлы), где происходит их дальнейшее созревание, презентация антигена, пролиферация и дифференцировка в плазматические клетки и В-клетки памяти.
Маркеры В-клеток
Характерной особенностью В-клеток является наличие поверхностных мембраносвязанных антител, относящихся к классам IgM и IgD. В комплексе с другими поверхностными молекулами иммуноглобулины формируют антигенраспознающий рецептивный комплекс, ответственный за узнавание антигена. Также на поверхности В-лимфоцитов расположены антигены МНС класса II, важные для взаимодействия с Т-клетками, также на некоторых клонах В-лимфоцитов присутствует маркер CD5, общий с Т-клетками. Рецепторы компонентов комплемента C3b (Cr1, CD35) и C3d (Cr2, CD21) играют определённую роль в активации В-клеток. Следует отметить, что маркеры CD19, CD20 и CD22 используются для идентификации В-лимфоцитов. Также на поверхности В-лимфоцитов обнаружены Fc рецепторы.
Дифференциация В-лимфоцитов
Лимфоциты происходят от плюрипотентных стволовых клеток, дающих также начало всем клеткам крови. Дифференциация стволовых клеток крови по эритроидному, миелоидному либо лимфоидному пути зависит от микроокружения (в случае птиц дифференциация стволовых клеток в В-лимфоциты происходит в фабрициевой сумке, у млекопитающих в костном мозге, где также происходит дифференциация по миелоидному и эритроидному пути). Дифференциация В-лимфоцитов условно делится на две стадии — антигеннезависимую (в которую происходит перестройка генов иммуноглобулинов и их экспрессия) и антигензависимую (при которой происходит активация, пролиферация и дифференциация в плазматические клетки).
- Пре-В-Клетки-предшественники не синтезируют тяжёлых и лёгких цепей, содержат зародышевые H и L гены, но содержат антигенный маркер, общий с зрелыми пре-В-клетками.
- Ранние пре-В-клетки — D-J перестройки в Н генах.
- Поздние пре-В-клетки — V-DJ перестройки в Н генах.
- Большие пре-В-клетки Н-гены VDJ-перестроены; в цитоплазме имеются тяжёлые цепи класса μ.
- Малые пре-В-клетки — V-J перестройки в L генах; в цитоплазме имеются тяжёлые цепи класса μ.
- Малые незрелые В-клетки — L гены VJ-перестроены; синтезируют Н и L- цепи; на мембране расположены иммуноглобулины.
- Зрелые В-клетки- начало синтеза IgD.
В-клетки клетки поступают из костного мозга во вторичные лимфоидные органы (селезёнку и лимфатические узлы), где происходит их дальнейшее созревание, антиген-презентирование, пролиферация и дифференциация в плазматические клетки и В-клетки памяти.
В-лимфоциты. В-клетки. Созревание В-лимфоцитов. Продолжительность жизни B-лимфоцитов. Маркеры В-клеток ( В-лимфоцитов ).
Известны субпопуляции В-клеток: предшественники антителообразующих (плазматических) клеток и В-клетки памяти (эффекторы вторичных иммунных реакций). Доминирующую субпопуляцию составляют предшественники антителообразующих клеток, дифференцирующиеся после антигенной стимуляции в плазматические клетки (плазмоциты), синтезирующие Ig.
Маркеры В-клеток ( В-лимфоцитов )
Основные маркёры В-лимфоцитов — мембранные Ig при этом клетки одного клона (быстро формирующегося в результате серии последовательных делений потом-ства одной В-клетки ) экспрессируют молекулы Ig, специфически связывающие только один эпитоп Аг. Такие клетки синтезируют моноклональные AT, способные распознавать и связывать только один Аг. Аг-связывающий участок мембранного Ig В-лимфоцита играет роль клеточного Аг-распознающего рецептора.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Активация В-клеток
Антигенпрезентирующие клетки (макрофаги, клетки Купфера, фолликулярные дендритные клетки, интердигитальные дендритные клетки и т. д.) вскоре после процессирования патогена выносят эпитопы на поверхность клетки при помощи белков МНС II, делая их доступными для Т-клеток. Т-хелпер при помощи Т-клеточного рецептора распознаёт комплекс эпитоп—МНС II. Активированный Т-хелпер выделяет цитокины, усиливающие антигенпрезентирующую функцию, а также цитокины, активирующие В-лимфоцит — индукторы активации и пролиферации. В-лимфоциты присоединяются при помощи мембраносвязанных антител, выступающих в роли рецепторов, к "своему" антигену и в зависимости от получаемых от Т-хелпера сигналов пролиферируют и дифференцируются в плазматическую клетку, синтезирующую антитела, либо перерождаются в В-клетки памяти. При этом от качества и количества антигена будет зависеть исход взаимодействия в данной трёхклеточной системе. Описанный механизм справедлив для полипептидных антигенов, относительно неустойчивых к фагоцитарному процессингу — т. н. тимусзависимых антигенов. Для тимуснезависимых антигенов (обладающих высокой полимерностью с часто повторяющимися эпитопами, относительно устойчивых к фагоцитарному перевариванию и обладающих свойствами митогена) участия Т-хелпера не требуется — активация и пролиферация В-лимфоцитов происходит за счёт собственной митогенной активности антигена.
Активация В-клеток
Антиген-презентирующая клетка (макрофаги, клетки Купфера, фолликулярные дендритные клетки, интердигитальные дендритные клетки и т. д.) вскоре после переваривания патогена выносит эпитопы на поверхность клетки при помощи МНС I или II (в зависимости от природы антигена), делая их доступными для Т-клеток. Т-хелпер при помощи Т-клеточного рецептора распознаёт комплекс эпитоп-МНС. Активированный Т-хелпер выделяет цитокины, усиливающие антиген-презентирующую функцию, а также цитокины, активирующих В-лимфоцит - индукторы активации и пролиферации. В-лимфоциты присоединяются при помощи мембрано-связанных антител, выступающих в роли рецепторов, к "своему" антигену и в зависимости от получаемых от Т-хелпера сигналов пролиферируют и дифференцируются в плазматическую клетку, синтезирующую антитела, либо перерождается в В-клетку памяти. При этом от качества и количества антигена будет зависеть исход данной трёхклеточной системы взаимодействия. Данный механизм справедлив для полипептидных антигенов, относительно неустойчивых к фагоцитарному процессингу - т. н. тимус-зависимых антигенов. Для тимус-независимых антигенов (обладающих высокой полимерностью с часто повторяющимися эпитопами, относительно устойчивых к фагоцитарному перевариванию и обладающих свойствами митогена) участия Т-хелпера не требуется - активация В-лимфоцитов происходит по тимус-независимому пути, В-лимфоциты связываются с данными антигенами, а за счёт их собственной митогенной активности будет происходить пролиферация В-лимфоцитов и активация.
В-лимфоциты. Характеристика В-лимфоцитов. Клетки памяти.
Предшественники В-лимфоцитов из эмбриональной печени человека на 8—9-й неделе развития плода перемещаются в костный мозг, где они созревают под влиянием интерлейкина 7, который секретируют стромальные клетки.
В ранний период созревания в костном мозге формируется и фиксируется на поверхностной мембране В-лимфоцита иммуноглобулиновый антигенраспознающий рецептор. Незрелый В-лимфоцит уже несет мембранные В-клеточные рецепторы (IgM) определенной специфичности, т. е. способные распознать и связать конкретный антиген. При встрече незрелых В-лимфоцитов с аутоантигенами связывание последних с мембранными IgM ведет к остановке дифференцировки этих В-лимфоцитов, с последующей их гибелью путем апоптоза. На этой стадии погибают до 90 % незрелых В-клеток. Часть незрелых В-клеток после контакта с растворимым аутоанти-геном не погибают, а переходят в состояние анергии по отношению к данному антигену. Этим объясняется врожденное состояние иммунологической толерантности (неспособности ответить) на собственные аутоантигены.
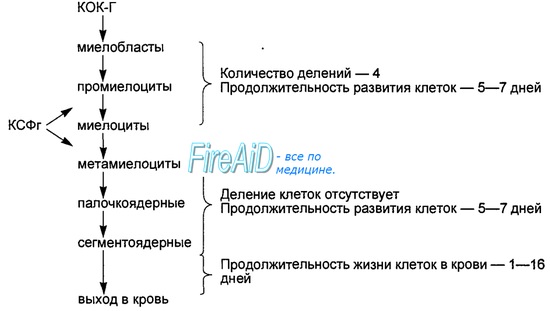
Выжившие в этом процессе отбора В-клетки по мере созревания покидают костный мозг, переходя в кровяное русло через венозные синусы, и поступают в периферические органы иммунной системы, где превращаются в зрелые В-лимфоциты. В кровяном русле В-лимфоциты включаются в рециркуляцию и заселяют В-зоны периферических органов и тканей иммунной системы.
Около 2 • 107 В-лимфоцитов покидают костный мозг ежедневно, что составляет 5—10 % общей популяции В-клеток периферической крови. Этого достаточно, чтобы полностью обновить популяцию периферических В-лимфоцитов за 4—5 дней. Среди сохранившихся и вступивших в рециркуляцию относительно долгоживущих В-лимфоцитов (с полупериодом жизни 1—2 мес) значительную часть составляют клетки памяти, способные быстро ответить пролиферацией, активацией, продукцией и секрецией специфических антител (иммуноглобулинов) на повторную встречу с тем же антигеном.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.

B-лимфоци́ты (B-клетки, от bursa fabricii птиц, где впервые были обнаружены) — функциональный тип лимфоцитов, играющих важную роль в обеспечении гуморального иммунитета.
У эмбрионов человека и других млекопитающих B-лимфоциты образуются в печени и костном мозге из стволовых клеток, а у взрослых млекопитающих — в красном костном мозге.
При контакте с антигеном или стимуляции со стороны T-клеток некоторые B-лимфоциты трансформируются в плазматические клетки, способные к продукции антител. Другие активированные B-лимфоциты превращаются в B-клетки памяти (см. ниже).
Содержание
В-клетки


Экспрессия всеми В-клетками мембранных иммуноглобулинов позволяет осуществляться клональному отбору под действием антигена. При созревании, стимулировании антигеном и пролиферации существенно меняется набор маркеров В-клеток. По мере созревания В-клетки переключаются от синтеза IgM и IgD на синтез IgG, IgA, IgE (при этом у клеток сохраняется способность синтезировать также IgM и IgD — вплоть до трёх классов одновременно). При переключении синтеза изотипов антигенная специфичность антител сохраняется. Различают следущие типы зрелых В-лимфоцитов:
- Собственно В-клетки (ещё называемые «наивными» В-лимфоцитами) — неактивированные В-лимфоциты, не контактировавшие с антигеном. Не содержат тельца Голла, в цитоплазме рассеяны монорибосомы. Полиспецифичны и имеют слабое сродство к многим антигенам.
- В-клетки памяти — активированые В-лимфоциты, вновь перешедшие в стадию малых лимфоцитов в результате кооперации с Т-клетками. Являются долгоживущим клоном В-клеток, обеспечивают быстрый иммунный ответ и выработку большого количества иммуноглобулинов при повторном введении того же антигена. Названы клетками памяти, так как позволяют иммунной системе «помнить» антиген на протяжении многих лет после прекращения его действия. В-клетки памяти обеспечивают долговременный иммунитет.
- Плазматические клетки — являются последним этапом дифференцировки активированных антигеном В-клеток. В отличие от остальных В-клеток несут мало мембранных антител и способны секретировать растворимые антитела. Являются большими клетками с эксцентрично расположенным ядром и развитым синтетическим аппаратом - шероховатый эндоплазматический ретикулум занимает почти всю цитоплазму, также развит и аппарат Гольджи. Являются короткоживущими клетками (2-3 дня) и быстро элиминируются при отсутствии антигена, вызвавшего иммунный ответ.
Клетки В-1 и В-2
Выделяют две субпопуляции В-клеток: В-1 и B-2. Субпопуляцию В-2 состовляют обычные В-лимфоциты, к которым относится всё сказанное выше. В-1 — это относительно небольшая группа В-клеток, обнаруживаемая у человека и мышей. Они могут составлять около 5% от общей популяции B-клеток. Такие клетки появляются в течение эмбрионального периода. На своей поверхности они экспрессируют IgM и небольшое количество (или вовсе не экспрессируют) IgD. Маркером этих клеток является CD5. Однако он не является обязательным компонентом клеточной поверхности. В эмбриональном периоде В1-клетки появляются из стволовых клеток костного мозга. В течение жизни пул B-1-лимфоцитов поддерживается за счёт активности специализированных клеток–предшественников и не пополняется за счёт клеток, происходящих из костного мозга. Клетка–предшественница отселяется из кроветворной ткани на свою анатомическую нишу — в брюшную и плевральную полости — ещё в эмбриональном периоде. Итак, место обитания B-1-лимфоцитов — прибарьерные полости.
B-1-лимфоциты значительно отличаются от B-2-лимфоцитов по антигенной специфичности продуцируемых антител. Антитела, синтезированные B-1-лимфоцитами, не имеют значительного разнообразия вариабельных участков молекул иммуноглобулинов, но, напротив, ограничены в репертуаре распознаваемых антигенов, и эти антигены — наиболее распространённые соединения клеточных стенок бактерий. Все B-1-лимфоциты — как бы один не слишком специализированный, но определённо ориентированный (антибактериальный) клон. Антитела, продуцируемые B-1-лимфоцитами, почти исключительно IgM, переключение классов иммуноглобулинов в B-1-лимфоцитах не «предусмотрено». Таким образом, B-1-лимфоциты — «отряд» противобактериальных «пограничников» в прибарьерных полостях, предназначенных для быстрой реакции на «просачивающиеся» через барьеры инфекционные микроорганизмы из числа широко распространённых. В сыворотке крови здорового человека преобладающая часть иммуноглобулинов — продукт синтеза как раз B-1-лимфоцитов, т.е. это относительно полиспецифичные иммуноглобулины антибактериального назначения.
B-лимфоци́ты (B-клетки, от bursa fabricii птиц, где впервые были обнаружены) — функциональный тип лимфоцитов, играющих важную роль в обеспечении гуморального иммунитета.
У эмбрионов человека и других млекопитающих B-лимфоциты образуются в печени и костном мозге из стволовых клеток, а у взрослых млекопитающих — в красном костном мозге.
При контакте с антигеном или стимуляции со стороны T-клеток некоторые B-лимфоциты трансформируются в плазматические клетки, способные к продукции антител. Другие активированные B-лимфоциты превращаются в B-клетки памяти.
Содержание
Роль В-лимфоцитов в презентации антигена
В-клетки способны поглощать свои мембранные иммуноглобулины вместе со связанным ими антигеном, а затем презентировать фрагменты антигена в комплексе с молекулами МНС класса II. При низкой концентрации антигена и при вторичном иммунном ответе В-клетки могут выполнять роль основных антигенпрезентирующих клеток.
Маркеры В-клеток
Характерной особенностью В-клеток является наличие поверхностных мембрано-связанных антител, относящихся к классам IgM и IgD. В комплексе с с другими поверхностными молекулами иммуноглобулины формируют антиген-распознающий рецептивный комплекс, ответственный за узнавание антигена. Также на поверхности В-лимфоцитов расположены антигены МНС класса II, важные в кооперации с Т-клетками, также на некоторых клонах В- лимфоцитов присутствует CD5 маркер, общий с Т-клетками. Рецепторы C3b компонента комплемента(Cr1, CD35) и C3d (Cr2,CD21) имеют определённую роль в активации В-клеток. Следует отметить, что маркеры CD19, CD20 и CD22 используются для идентификации В-лимфоцитов. Также на поверхности В-лимфоцитов обнаружены Fc-рецепторы.
Читайте также: